The Scientist and Engineer's Guide to
Digital Signal Processing
By Steven W. Smith, Ph.D.
Book Search

Table of contents
- 1: The Breadth and Depth of DSP
- 2: Statistics, Probability and Noise
- 3: ADC and DAC
- 4: DSP Software
- 5: Linear Systems
- 6: Convolution
- 7: Properties of Convolution
- 8: The Discrete Fourier Transform
- 9: Applications of the DFT
- 10: Fourier Transform Properties
- 11: Fourier Transform Pairs
- 12: The Fast Fourier Transform
- 13: Continuous Signal Processing
- 14: Introduction to Digital Filters
- 15: Moving Average Filters
- 16: Windowed-Sinc Filters
- 17: Custom Filters
- 18: FFT Convolution
- 19: Recursive Filters
- 20: Chebyshev Filters
- 21: Filter Comparison
- 22: Audio Processing
- 23: Image Formation & Display
- 24: Linear Image Processing
- 25: Special Imaging Techniques
- 26: Neural Networks (and more!)
- 27: Data Compression
- 28: Digital Signal Processors
- 29: Getting Started with DSPs
- 30: Complex Numbers
- 31: The Complex Fourier Transform
- 32: The Laplace Transform
- 33: The z-Transform
- 34: Explaining Benford's Law
How to order your own hardcover copy
Wouldn't you rather have a bound book instead of 640 loose pages?Your laser printer will thank you!
Order from Amazon.com.
Chapter 22: Audio Processing
The human ear is an exceedingly complex organ. To make matters even more difficult, the information from two ears is combined in a perplexing neural network, the human brain. Keep in mind that the following is only a brief overview; there are many subtle effects and poorly understood phenomena related to human hearing.
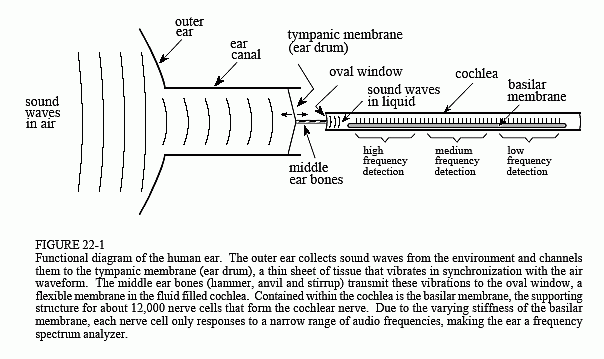
Figure 22-1 illustrates the major structures and processes that comprise the human ear. The outer ear is composed of two parts, the visible flap of skin and cartilage attached to the side of the head, and the ear canal, a tube about 0.5 cm in diameter extending about 3 cm into the head. These structures direct environmental sounds to the sensitive middle and inner ear organs located safely inside of the skull bones. Stretched across the end of the ear canal is a thin sheet of tissue called the tympanic membrane or ear drum. Sound waves striking the tympanic membrane cause it to vibrate. The middle ear is a set of small bones that transfer this vibration to the cochlea (inner ear) where it is converted to neural impulses. The cochlea is a liquid filled tube roughly 2 mm in diameter and 3 cm in length. Although shown straight in Fig. 22-1, the cochlea is curled up and looks like a small snail shell. In fact, cochlea is derived from the Greek word for snail.
When a sound wave tries to pass from air into liquid, only a small fraction of the sound is transmitted through the interface, while the remainder of the energy is reflected. This is because air has a low mechanical impedance (low acoustic pressure and high particle velocity resulting from low density and high compressibility), while liquid has a high mechanical impedance. In less technical terms, it requires more effort to wave your hand in water than it does to wave it in air. This difference in mechanical impedance results in most of the sound being reflected at an air/liquid interface.
The middle ear is an impedance matching network that increases the fraction of sound energy entering the liquid of the inner ear. For example, fish do not have an ear drum or middle ear, because they have no need to hear in air. Most of the impedance conversion results from the difference in area between the ear drum (receiving sound from the air) and the oval window (transmitting sound into the liquid, see Fig. 22-1). The ear drum has an area of about 60 (mm)2, while the oval window has an area of roughly 4 (mm)2. Since pressure is equal to force divided by area, this difference in area increases the sound wave pressure by about 15 times.
Contained within the cochlea is the basilar membrane, the supporting structure for about 12,000 sensory cells forming the cochlear nerve. The basilar membrane is stiffest near the oval window, and becomes more flexible toward the opposite end, allowing it to act as a frequency spectrum analyzer. When exposed to a high frequency signal, the basilar membrane resonates where it is stiff, resulting in the excitation of nerve cells close to the oval window. Likewise, low frequency sounds excite nerve cells at the far end of the basilar membrane. This makes specific fibers in the cochlear nerve respond to specific frequencies. This organization is called the place principle, and is preserved throughout the auditory pathway into the brain.
Another information encoding scheme is also used in human hearing, called the volley principle. Nerve cells transmit information by generating brief electrical pulses called action potentials. A nerve cell on the basilar membrane can encode audio information by producing an action potential in response to each cycle of the vibration. For example, a 200 hertz sound wave can be represented by a neuron producing 200 action potentials per second. However, this only works at frequencies below about 500 hertz, the maximum rate that neurons can produce action potentials. The human ear overcomes this problem by allowing several nerve cells to take turns performing this single task. For example, a 3000 hertz tone might be represented by ten nerve cells alternately firing at 300 times per second. This extends the range of the volley principle to about 4 kHz, above which the place principle is exclusively used.
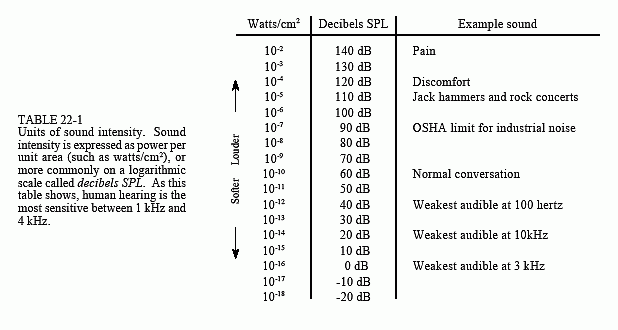
Table 22-1 shows the relationship between sound intensity and perceived loudness. It is common to express sound intensity on a logarithmic scale, called decibel SPL (Sound Power Level). On this scale, 0 dB SPL is a sound wave power of 10-16 watts/cm2, about the weakest sound detectable by the human ear. Normal speech is at about 60 dB SPL, while painful damage to the ear occurs at about 140 dB SPL.
The difference between the loudest and faintest sounds that humans can hear is about 120 dB, a range of one-million in amplitude. Listeners can detect a change in loudness when the signal is altered by about 1 dB (a 12% change in amplitude). In other words, there are only about 120 levels of loudness that can be perceived from the faintest whisper to the loudest thunder. The sensitivity of the ear is amazing; when listening to very weak sounds, the ear drum vibrates less than the diameter of a single molecule!
The perception of loudness relates roughly to the sound power to an exponent of 1/3. For example, if you increase the sound power by a factor of ten, listeners will report that the loudness has increased by a factor of about two (101/3 ≈ 2). This is a major problem for eliminating undesirable environmental sounds, for instance, the beefed-up stereo in the next door apartment. Suppose you diligently cover 99% of your wall with a perfect soundproof material, missing only 1% of the surface area due to doors, corners, vents, etc. Even though the sound power has been reduced to only 1% of its former value, the perceived loudness has only dropped to about 0.011/3 ≈ 0.2, or 20%.
The range of human hearing is generally considered to be 20 Hz to 20 kHz, but it is far more sensitive to sounds between 1 kHz and 4 kHz. For example, listeners can detect sounds as low as 0 dB SPL at 3 kHz, but require 40 dB SPL at 100 hertz (an amplitude increase of 100). Listeners can tell that two tones are different if their frequencies differ by more than about 0.3% at 3 kHz. This increases to 3% at 100 hertz. For comparison, adjacent keys on a piano differ by about 6% in frequency.
The primary advantage of having two ears is the ability to identify the direction of the sound. Human listeners can detect the difference between two sound sources that are placed as little as three degrees apart, about the width of a person at 10 meters. This directional information is obtained in two separate ways. First, frequencies above about 1 kHz are strongly shadowed by the head. In other words, the ear nearest the sound receives a stronger signal than the ear on the opposite side of the head. The second clue to directionality is that the ear on the far side of the head hears the sound slightly later than the near ear, due to its greater distance from the source. Based on a typical head size (about 22 cm) and the speed of sound (about 340 meters per second), an angular discrimination of three degrees requires a timing precision of about 30 microseconds. Since this timing requires the volley principle, this clue to directionality is predominately used for sounds less than about 1 kHz.
Both these sources of directional information are greatly aided by the ability to turn the head and observe the change in the signals. An interesting sensation occurs when a listener is presented with exactly the same sounds to both ears, such as listening to monaural sound through headphones. The brain concludes that the sound is coming from the center of the listener's head!
While human hearing can determine the direction a sound is from, it does poorly in identifying the distance to the sound source. This is because there are few clues available in a sound wave that can provide this information. Human hearing weakly perceives that high frequency sounds are nearby, while low frequency sounds are distant. This is because sound waves dissipate their higher frequencies as they propagate long distances. Echo content is another weak clue to distance, providing a perception of the room size. For example, sounds in a large auditorium will contain echoes at about 100 millisecond intervals, while 10 milliseconds is typical for a small office. Some species have solved this ranging problem by using active sonar. For example, bats and dolphins produce clicks and squeaks that reflect from nearby objects. By measuring the interval between transmission and echo, these animals can locate objects with about 1 cm resolution. Experiments have shown that some humans, particularly the blind, can also use active echo localization to a small extent.